20_4.jpg
20_1.jpg
20_2.jpg
20_3.jpg
20_5.jpg
20_6.jpg
20_7.jpg
20_8.jpg
EGSA Grad Student Handbook
PDF document, 686.6 KB
Handbook prepared by students of the Entomological Graduate Student Association (EGSA)
Julie Todd CV
Microsoft Word Document, 108.0 KB
2019
Student Handbook
elena-gratton.png
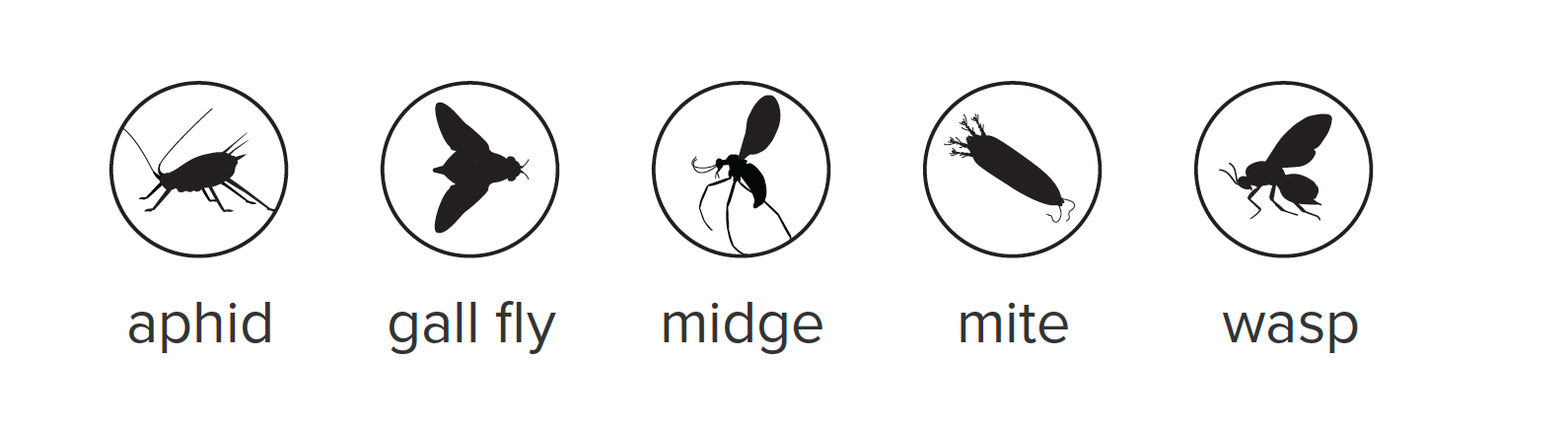
wasp-galls-insect-key.png
Archbold-9.jpg
IMGP3048 - Copie.jpg
2023 Center for Pollinator Research | Bee Championship
PDF document, 9.2 MB
Create your bracket- who is your 2023 Bee of the Year?
adam-scherr1.jpg
adam-scherr2.png
wordcloud2023.gif
New_Insect_ID_Request_fillable-2024.pdf
PDF document, 198.4 KB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}